Hypothesis 1
- Changes in mean behavior -
- Exploration 1 (Swimming distance) -
The results imply, combined with the Stage and Stage:Sex effect, that food as an environmental cue fostering changes in exploratory behavior did not take effect until later in life. Subadults in both treatments did increase exploration in a similar manner but in the adult stage food effect became evident. Edenbrow & Croft however observed an adverse effect in mangrove killifish (Kryptolebias marmoratus). In their study, fish reared under low-food conditions expressed a reduced exploratory behaviour when compared to a control group. According to them, low-food fish reduced exploratory behaviour in order to counter size selective mortality (Edenbrow & Croft 2013; Sogard 1997). They were not significantly shyer than conspecifics, their reduced boldness trajectories yet indicate evidence of size selective risk sensitivity (Edenbrow & Croft 2013). Results from my study differed possibly due to the fact of removing predation risk all together. A constant absence of predatory cues could have enhanced the urge to increase foraging and exploration without fearing adverse effects like an increase in mortality (Fawcett & Frankenhuis 2015; Panchanathan & Frankenhuis 2016).
- Exploration 2 (Moving time) -
I would conclude that the increased exploratory behavior expressed by increased moving time for low-food males compared to high-food males was a response to scarcity in food (Fawcett & Frankenhuis 2015; Nislow et al. 1998). For high-food females it was potentially their higher overall energy demand that led to an increase in exploration compared to same treatment males (Magurran & Garcia 2005; Rljnsdorp & Ibelings 1989). Low-food females showed the same trend but it was non-significant as mentioned before, possibly as a measure to conserve energy by moving less (Orpwood et al. 2006). Food thus had a very specific influence on time spent moving with different effects depending on sex and treatment. Food effects may be intertwined with physiology. Sexes in dimorphic species generally express differences in aspects like aggression, foraging and fitness (Magurran & Garcia 2005; Sogard & Olla 1996). This should be considered when trying to estimate the influential value of food in combination with sex.
- Shyness (Time spent in outer area) -
First of all, there was a stage-effect. Subadult fish became shyer than they were in their juvenile stage and later became less shy, i.e. bolder, again as adults. Brown & Braithwaite observed a higher risk-taking behaviour in juvenile fish compared to larger and older ones (Brachyraphis episcopi). They explained said behaviour by a metabolic hypothesis, considering the higher urge for juvenile fish to feed and grow (Brown & Braithwaite 2004). For juveniles, the need to feed and grow might have reduced expressed shyness (Ballew et al. 2017; Biro et al. 2005). As for subadult fish scientific literature states the subadult stage as the most crucial for many species in terms of maturation and potential future reproductive success. An increase in shyness during that stage can be seen as a form of asset protection (Reinhardt & Healey 1999). Therefore, subadult Heterandria formosa preferred the safer outer areas to reduce risks by being shyer than in previous and later stages.
The results of the experiment show that for a Stage:Food effect, high-food subadults were significantly shyer than low-food subadults, before both fish in both treatments became less shy again upon adulthood. High-food conditions allow an animal to be shyer since the need to increase risk-taking to increase foraging is not as pressing as under low-food conditions (Brown & Braithwaite 2004; Conrad et al. 2011).
A treatment effect could not be generalized but food and its effect on shyness seemed to have been highly stage dependent, fostering fish to be less shy in their subadult stage when facing scarceness of food and at the same time to be shyer under high-food conditions.
The results of the experiment show that for a Stage:Food effect, high-food subadults were significantly shyer than low-food subadults, before both fish in both treatments became less shy again upon adulthood. High-food conditions allow an animal to be shyer since the need to increase risk-taking to increase foraging is not as pressing as under low-food conditions (Brown & Braithwaite 2004; Conrad et al. 2011).
A treatment effect could not be generalized but food and its effect on shyness seemed to have been highly stage dependent, fostering fish to be less shy in their subadult stage when facing scarceness of food and at the same time to be shyer under high-food conditions.
- Activity (Actual velocity) -
There was a distinct increase in swimming speed across stages. When they were swimming, fish swam faster on average during later life-stages.
The increase in swimming speed for adult females was mostly attributed to low-food female adults whereas high-food female adults decreased in swimming speed upon maturation which hints at a foraging motivation for low-food fish to increase activity (Brown & Braithwaite 2004; Smith & Blumstein 2008). However, this was only a trend and food was not in any way a significant factor for differences in actual velocity. As a consequence, there was no detectable food treatment effect and variations in actual velocity over ontogeny should be attributed to physiological changes and size at age effects (Jones et al. 1974; Maguit et al. 2010).
The increase in swimming speed for adult females was mostly attributed to low-food female adults whereas high-food female adults decreased in swimming speed upon maturation which hints at a foraging motivation for low-food fish to increase activity (Brown & Braithwaite 2004; Smith & Blumstein 2008). However, this was only a trend and food was not in any way a significant factor for differences in actual velocity. As a consequence, there was no detectable food treatment effect and variations in actual velocity over ontogeny should be attributed to physiological changes and size at age effects (Jones et al. 1974; Maguit et al. 2010).
Key points
- At a glance -
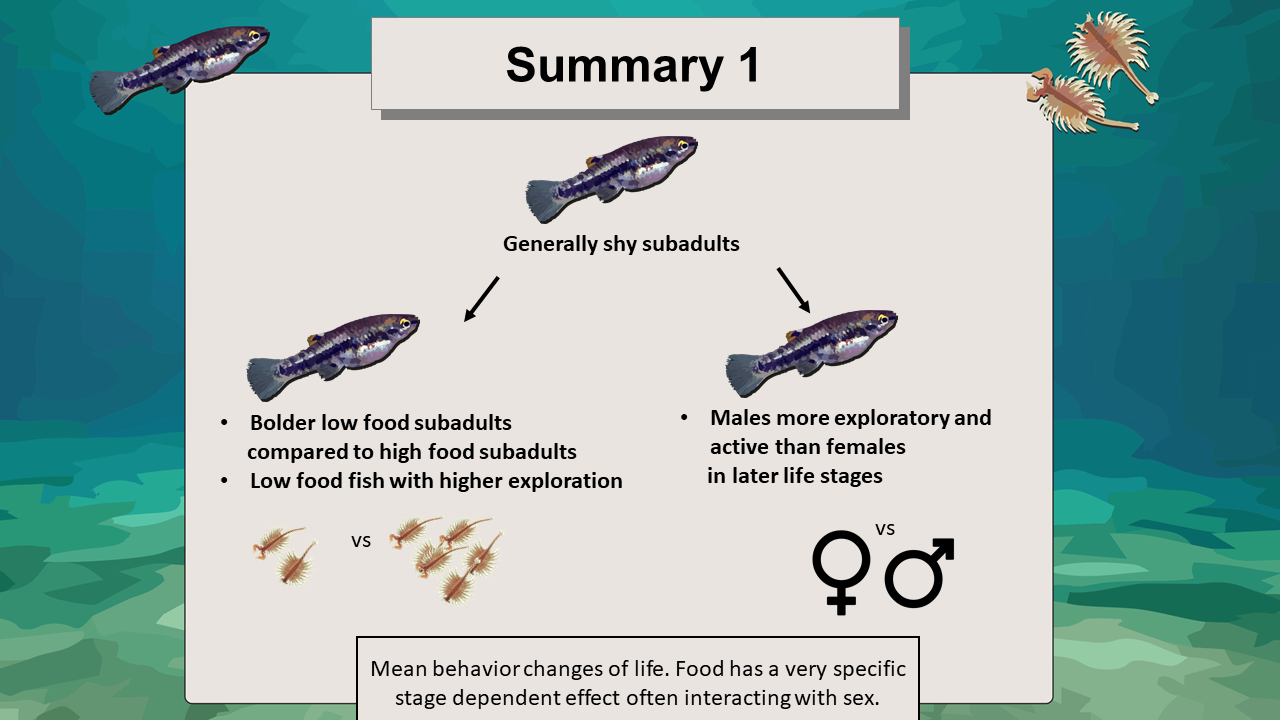
Figure 33. Main findings in regards to Stage, Sex and Food effects on mean behavior.
Hypothesis 2
- Among and within variation -
Consolidation of behavior with age in my case was attributed due to higher among-individual variation of older fish. the test fish became more distinct from one another with age but did not change in their individual plasticity during their life-stages as assessed during the four consecutive behavioural trials.
The question remains now why fish developed behavioral differences when compared to each other and consolidate those throughout life, independent of sex and food? In this regard, an experiential effect could be a possible reason but only to some extent, as the conditions for each fish did not change significantly over ontogeny (Bell et al 2009; Conrad et al. 2011). As one of the newest study available tackling that topic, Bierbach et al. found that individual variation occurred even in clonal generations of the Amazon molly (Poecilia formosa), despite their clonal nature and near identical rearing conditions (Bierbach et al 2017). According to Bierbach et al., it could be that “individuality might be an inevitable and potentially unpredictable outcome of development” (Bierbach et al. 2017). They argue that so called “minute experiential and environmental experiences” could shape behavioral differentiation more than previously thought (Bierbach et al. 2017).
As for food, it only had an influence on the second exploratory variable (moving time). Low-food fish had a significantly lower among-individual variation and repeatability than high-food fish. Among-individual differences and repeatability changed for low-food fish because under environmental food stress they were partially forced to a more uniform behavior. Low food fish varied less among each other because they all had to explore to compensate the low-food conditions. In contrast high-food fish did not have that limitation by having always enough food and developing without constraints in an individual direction (Conrad et al. 2011; Fawcett & Frankenhuis 2015). Note however that those findings are, only implications, since the food effect was only detectable for 1 out of 4 measured traits but nonetheless interesting under the aspect of further, more extensive studies.
The question remains now why fish developed behavioral differences when compared to each other and consolidate those throughout life, independent of sex and food? In this regard, an experiential effect could be a possible reason but only to some extent, as the conditions for each fish did not change significantly over ontogeny (Bell et al 2009; Conrad et al. 2011). As one of the newest study available tackling that topic, Bierbach et al. found that individual variation occurred even in clonal generations of the Amazon molly (Poecilia formosa), despite their clonal nature and near identical rearing conditions (Bierbach et al 2017). According to Bierbach et al., it could be that “individuality might be an inevitable and potentially unpredictable outcome of development” (Bierbach et al. 2017). They argue that so called “minute experiential and environmental experiences” could shape behavioral differentiation more than previously thought (Bierbach et al. 2017).
As for food, it only had an influence on the second exploratory variable (moving time). Low-food fish had a significantly lower among-individual variation and repeatability than high-food fish. Among-individual differences and repeatability changed for low-food fish because under environmental food stress they were partially forced to a more uniform behavior. Low food fish varied less among each other because they all had to explore to compensate the low-food conditions. In contrast high-food fish did not have that limitation by having always enough food and developing without constraints in an individual direction (Conrad et al. 2011; Fawcett & Frankenhuis 2015). Note however that those findings are, only implications, since the food effect was only detectable for 1 out of 4 measured traits but nonetheless interesting under the aspect of further, more extensive studies.
Summary and implications
- Main takeaway -
With age individuals became more and more distinct from each other in their behavior. Food only had limited influence on the expression of behavioral types as well as their development over ontogeny. However, when present, food seemed to affect behaviour during certain life-stages in a typical way. Scarcity in food, seemed to have led to a more constrained and uniform exploratory behaviour in Heterandria formosa when compared to high-food fish.
The measured mean behavior changed across life-stages. This was likely due to different energy requirements in each life-stage and the varying physiology of the fish across life -stages and between sexes (Conrad et al. 2011; Fawcett & Frankenhuis 2015). If food had an effect, it became evident through interactions with stage or sex. Those interactions and their effects suggest that the food shortage led to higher boldness and exploration to increase foraging (Conrad et al. 2011; Mittelbach et al. 2014).
Not all hypotheses could have been accepted and the results were sometimes inconsistent. This may be due to the selection of the behavioral variables measured. Especially the two variables potentially assessing exploratory behaviour but showing different results give room for improvement. While I based the variable selection and the interpretation on an extensive literature it may be that these selections and interpretations were not correct for the model system (despite that Heterandria formosa is, as many other fish model species like Guppies or Mollies, a member of the family of the Poecillidae).
Despite potential improvements and constraints, it remains that in contrast to many studies the study assessed behavioral changes under different food conditions and covered a full life history. The interesting and comprehensive results consequently underline the advantages of longitudinal studies and their importance for the scientific field of behavioral ecology.
The measured mean behavior changed across life-stages. This was likely due to different energy requirements in each life-stage and the varying physiology of the fish across life -stages and between sexes (Conrad et al. 2011; Fawcett & Frankenhuis 2015). If food had an effect, it became evident through interactions with stage or sex. Those interactions and their effects suggest that the food shortage led to higher boldness and exploration to increase foraging (Conrad et al. 2011; Mittelbach et al. 2014).
Not all hypotheses could have been accepted and the results were sometimes inconsistent. This may be due to the selection of the behavioral variables measured. Especially the two variables potentially assessing exploratory behaviour but showing different results give room for improvement. While I based the variable selection and the interpretation on an extensive literature it may be that these selections and interpretations were not correct for the model system (despite that Heterandria formosa is, as many other fish model species like Guppies or Mollies, a member of the family of the Poecillidae).
Despite potential improvements and constraints, it remains that in contrast to many studies the study assessed behavioral changes under different food conditions and covered a full life history. The interesting and comprehensive results consequently underline the advantages of longitudinal studies and their importance for the scientific field of behavioral ecology.
Key points
- At a glance -
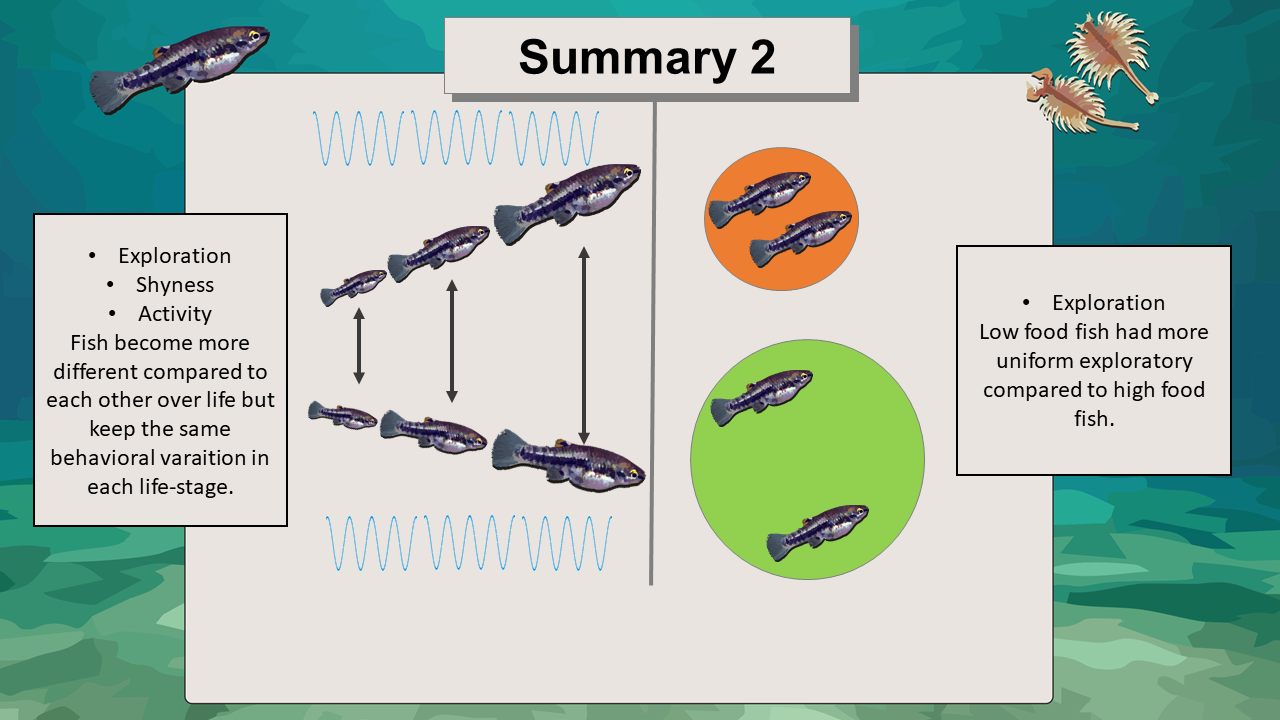
Figure 34. Main findings summary for repeatability, within and among individual variation in fish behavior.
